Evolution Basics: From Primate to Human
For many years, it was unclear if humans were more closely related to chimpanzees or to gorillas, but full genome sequences allowed us to resolve the issue.
In the last post in this series, we examined the earliest-known primates. This lineage would continue to expand and diversify, ultimately giving rise to a wide range of forms. Included in this diversity is what we now recognize as New World monkeys, Old World monkeys, lesser apes, and our own group, the great apes (hominids). Crown-group hominids include orangutans, gorillas, chimpanzees, bonobos, and humans; their last common ancestral population; and all extinct species descended from that common ancestral population. Present-day great apes are greatly reduced in species diversity compared to the past – in other words, they are relict populations, surviving in isolated, fragmented habitats. Humans are the sole exception to this pattern in terms of population size, but we too are the sole surviving lineage among our closest relatives, as we shall see.
Full genome sequences are now available for most living great ape species, including humans, chimpanzees, gorillas, and orangutans. These sequences have allowed us to determine that the lineages of these four species branched from each other in the following pattern: humans are most closely related to chimpanzees, then to gorillas, and then to orangutans:
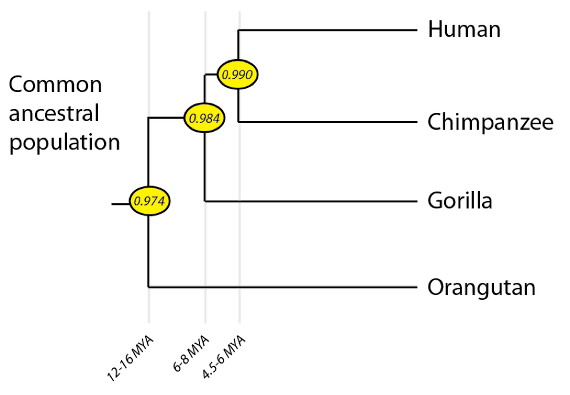
A phylogeny of present-day crown-group hominids (great apes). Numbers in yellow ovals at the branch points indicate the percent DNA identity with humans (excluding deletion and insertion mutations). Genome identity values and speciation times are from Locke et al., 2011.
For many years, it was unclear if humans were more closely related to chimpanzees or to gorillas, but full genome sequences allowed us to resolve the issue. One reason for the (now solved) controversy was that the gorilla and chimpanzee lineages branched off our own lineage in relatively quick succession, and did so with quite large population sizes. As you may recall from an earlier post in this series, rapid branching with large populations can lead to an effect known as incomplete lineage sorting, where some of our own genome is more closely related to gorillas than to chimpanzees. Due to this effect, phylogenies for individual genes sometimes produce a ((human, gorilla) chimpanzee) pattern rather than the overall ((human, chimpanzee) gorilla) pattern. As expected, there are even a tiny number of genes in the human genome that are more closely related to the orangutan version of the gene, as I discussed back in 2011 when the orangutan genome was published. Full-genome sequence comparisons between living great apes also fit the expected pattern: orangutans and humans have sequences 97.4% identical; humans and gorillas, 98.4% identical; and humans and chimpanzees, 99.0% identical. (Note, this value does not include insertion/deletion mutations – so-called “indels” – if it did, the human-chimpanzee value would be ~95%, and the other comparisons would follow the same pattern of decreasing identity).
As an aside, the striking amount of genetic identity between humans and other great apes is sometimes puzzling to non-biologists. How is it that species that are so genetically similar be so markedly different? While we will explore this in more detail later in this series, the brief answer is that, for a complex organism, small changes at the DNA level can bring about quite dramatic changes in form and behavior – and these small changes are of the sort that are readily accessible to evolutionary mechanisms that shift allele frequencies in populations over time.
From hominid to hominin
Having surveyed the hominid crown group (orangutans, gorillas, chimpanzees and humans, their last common ancestral population, and all descendant species of that population), we are now prepared to examine our own branch within it – species more closely related to us than to chimpanzees. Such species are known as hominins. Though humans are the only surviving hominin lineage, there was once a wide diversity of hominin species on the planet, some of which lived alongside early humans. One interesting fact about our own lineage is that our “branch” emerges from a rather “bushy” phylogeny. There are many hominin forms in the fossil record, and teasing out their precise relatedness to one another is a challenging exercise (for more detail, see the series by anthropologist James Kidder in “For further reading” below). Forms for which DNA sequence is available are easy to place in a phylogeny, but those known only from fossil remains are more difficult to place. Given our forgoing discussion of stem-group and crown-group species, however, we are now prepared to appreciate these fossil hominins for what they are: stem-group species on our own branch, with some species possibly ancestral to our own, or located very close to the branch points with our lineage:
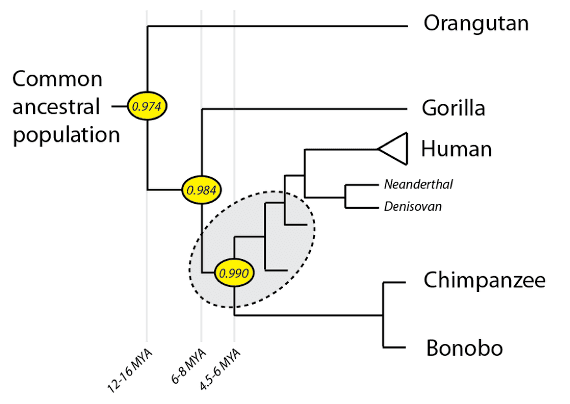
A phylogeny of crown-group hominids (great apes). Humans, together with extinct species more closely related to humans than to chimpanzees, comprise a subset of hominids known as hominins. Extinct hominin groups such as Kenyanthropus, Paranthropus and Australopithecus are near root of the hominin tree (shaded in gray). Other groups such as Ardipithecus, Orrorin, and Sahelanthropus may be within the hominin tree or stem groups that branch off prior to the last common ancestor of humans and chimpanzees.
The area shaded in gray in the above phylogeny is the area of interest. Some forms in the fossil record can be readily identified as stem hominins (i.e. more closely related to us than chimpanzees) whereas others are more ambiguous: if indeed they branch off our lineage prior to our last common ancestral population with chimpanzees, they are in fact hominids, not stem-group hominins:
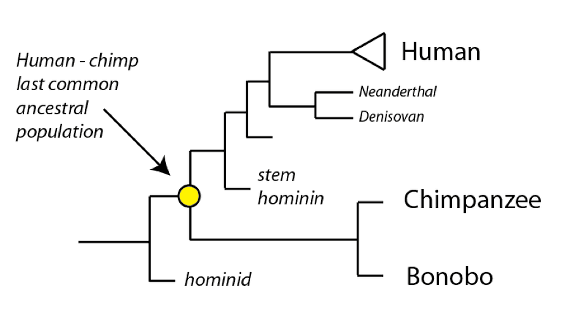
Some fossil species that may either be closely-related hominids or stem-group hominins are Sahelanthropus tchadensis, Orrorin tugenensis, and two ardipithecine species (Ardipithecus ramidus and Ardipithecus kadabba). Regardless of their precise placement on the tree, these species are close to the last common ancestral population of humans and chimpanzees, and provide some clues to what that ancestral species looked like, and the order in which we acquired our defining characteristics.
One key distinctive of the hominin lineage is bipedality: walking upright on two legs, rather than on all fours. There is some evidence for bipedal locomotion in Ardipithecus ramidus, Sahelanthropus, and Orrorin, though it is controversial within the field. The evidence is best in Ardipithecus ramidus, which appears to have been a facultative biped (able to walk upright as well as on all fours). Since these species all have small brain volumes (under 400 cubic centimeters), this evidence supports the view that bipedality evolved prior to the expansion in brain volume we observe in later, unambiguous hominins. As such it is probable that our last common ancestor with chimpanzees was an ape with a small brain case and facultative bipedality – traits that would then be shaped substantially in our lineage over time.
Previously, we examined species known from the fossil record that are near the last common ancestral population we share with our closest living relatives (chimpanzees). As we have discussed, species more closely related to us than to chimpanzees are known as hominins. As we will see, the lineage from which humans emerged is the sole remaining branch of what was once a diverse group of related species.
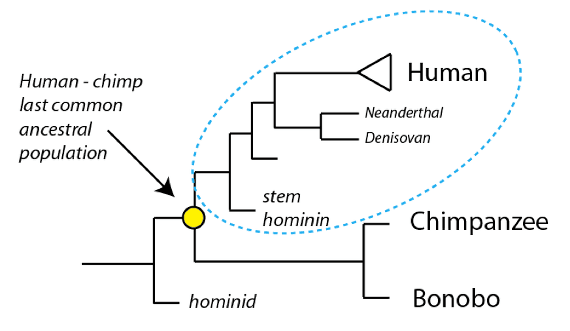
Hominins (outlined in blue) are species more closely related to humans than to chimpanzees. Humans are the sole surviving lineage of what was once a diverse group of hominin species. Hominins are nested within the hominids, which include all great apes, their last common ancestral population, and all descendant species of that population.
The paleontology of this group is interesting not only for its scientific aspects, but also for the history surrounding it. The intense interest surrounding our evolutionary relatives, preconceived ideas about how and where the transition from ape to human came about, and human pride and deception would all play a significant role in shaping the discovery and interpretation of the first hominin fossils.
Seeking the “missing link”
When Darwin published On the Origin of Species in 1859, and later The Descent of Man in 1871, scientific knowledge of the hominin fossil record was nearly non-existent. While the first Neanderthal bones had been discovered by the 1850s, an understanding of what these finds (and subsequent Neanderthal discoveries in the 1880s) represented would be decades in coming. Following on from Darwin’s work there was great interest in identifying transitional forms between humans and living apes. Neanderthals, however, were close enough in form to modern humans for scientists to doubt that they were a distinct species. What was expected, and sought, were hypothesized forms commonly viewed as “missing links” in a ladder-like progression from ape to human. The appreciation of the hominin group as a bushy tree rather than a ladder would elude scientists for some time, to say nothing of the general public.
It was in this context that Eugene Dubois would set out for Indonesia in the late 1880s to search for fossils linking humans and apes. (Despite Darwin concluding from the available evidence that human origins would be found in Africa, the early, largely preconceived consensus was that humans had originated in Asia.) Dubois was the first scientist to deliberately look for such fossils, and through great effort and personal hardship he was successful. Dubois discovered the first fossils of what would later be known as Homo erectus in Indonesia: a fossil he provocatively named Pithecanthropus erectus (“the ape-man that stands upright”). Unfortunately for Dubois, his finds were fragmentary – a few teeth, the cap of the skull, and a femur – and generally not well received by the scientific community of his day. Many suspected – given the small brain size of his specimen and the human-like leg bones – that Dubois’ “ape man” was nothing more than the coincidental discovery of human remains mixed with that of an extinct ape. This skepticism was fueled in part by the assumption at the time that it was brain evolution that had set humans apart from apes. The expected “missing link”, therefore, was expected to have an ape-like body and a more human-sized skull. It was not until decades later that Dubois’ interpretation of Pithecanthropus – that it indeed was a species with a human-like body and small ape-like brain – would be vindicated.
Dubois’ views would face a further setback in 1912, when a fossil discovery conveniently matching the expected set of intermediate features would be discovered: the infamous Piltdown Man. This “find” would later be shown to be a forgery constructed from a crushed modern human skull with the jaw of an orangutan, complete with teeth filed down to match expectations of the time for an intermediate between humans and living apes. As before, critics did raise doubts that perhaps the fossil was an amalgamation of an extinct ape jaw and a human skull, but these criticisms were largely silenced a few years later when a second skull matching the first was also “found” at a nearby site. Whereas chance might place the remains of two distinct creatures together once, it was highly unlikely it would do so twice – and so Piltdown gained acceptance for a time.
It would take considerable work to dethrone Piltdown as the “missing link” between humans and apes, but ultimately suspicion would grow to the point where it was exposed as a fraud. That work was the ongoing discovery of legitimate hominin remains – discoveries that increasingly singled out Piltdown Man as an oddity that did not fit within the growing picture of hominin evolution.
Enter the Australopithecines
One such discovery that would eventually contribute to exposing Piltdown was a second hominin species that combined human-like traits with a sub-human braincase volume. In 1924, anatomist Raymond Dart would examine fossils from Tuang, South Africa and discover a juvenile hominin with a braincase volume too large for an ape, but below that even of Homo erectus, precluding it from being human. As Dart would later recount in his memoirs,
I knew at a glance that what lay in my hands was no ordinary anthropoidal brain. Here in lime-consolidated sand was the replica of a brain three times as large as that of a baboon and considerably bigger than that of an adult chimpanzee. The startling image of the convolutions and furrows of the brain and the blood vessels of the skull were plainly visible.
It was not big enough for primitive man, but even for an ape it was a big bulging brain and, most important, the forebrain was so big and had grown so far backward that it completely covered the hindbrain.
Dart published his work in 1925 in the influential journal Nature under with the title “Australopithecus africanus: The Man-Ape of South Africa”. Like Dubois before him, however, Dart’s seminal discovery would meet a cold reception – including the opposition of prominent anthropologists convinced not only that human evolution was a brain-first affair, but also that human origins were in Asia rather than Africa. These critics noted that unless anadult member of the species could be found, that the juvenile Dart had described was likely to be a mere ape – and not a species intermediate between apes and humans. Once again, the shadow of Piltdown Man – with its “correct” ape-like teeth and human-like braincase – hung over this new discovery with its incongruous mix of human-like teeth and a sub-human braincase volume. Like Dubois before him, Dart would have to await further discoveries for the tide of scientific opinion to turn.
Next, we’ll explore how continued work in hominin paleontology provided a clearer picture of hominin evolution and eventually exposed Piltdown Man as a fraud.
We have discussed the seminal discovery of Australopithecine fossils by Raymond Dart and the cool reception they received by the scientific establishment of the time. In the 1920s, the epicenter of science was England, and the English had their “missing link” bridging humans and apes – the fraudulent Piltdown Man, with its then-expected ape-like body and a human-like brain. This erroneous “brain first, body second” view of human evolution meant that the true hominin fossils then known (Dart’s Australopithecus and Dubois’ Pithecanthropus (Homo) erectus) were “backwards”, with human-like bodies matched to inappropriately small brains. As we have seen, Dubois’ finds were widely thought to be chance assemblages of human skeletal remains with an ape skull; similarly, Dart’s interpretation was criticized based on his specimen being a juvenile. Without an adult form to examine, it was surmised that he had merely discovered a species closely related to modern apes.
Dart did gain some allies, however – and two in particular would assist him in slowly turning the tide of scientific opinion to his interpretation of the australopithecines as true hominins. The first was Robert Broom, a somewhat eccentric paleontologist and Fellow of the Royal Society who would work with additional australopithecine fossil discoveries in the 1930s. One such find was an exceptional fossil skull Broom would describe as Plesianthropus transvaalensis, a species distinct from Dart’s Australopithecus africanus. From its scientific name the fossil would soon acquire the moniker “Mrs. Ples”, an affectionate nickname that has stuck despite its later reclassification as an adult Australopithecus africanus rather than a separate australopithecine species. A few years later, however, Broom would discover a genuinely new hominin fossil – a species of stocky, muscular australopithecine he would name Australopithecus robustus (a species closely related to the australopithecines that was later renamed Paranthropus robustus). These additional finds greatly bolstered Dart’s position, since they too had the “body first” characteristics in contrast to Piltdown Man.
Dart’s second ally came in the person of Wilfrid Le Gros Clark, also a Fellow of the Royal Society, and well-respected by the scientific establishment. In 1947 Clark would write an influential paper on the australopithecines, chastise the establishment for passing judgment on them without examining them for themselves as he had done (as only a polite, yet scathing Englishman of the time could), and exonerate Dart and Broom in the process:
On the basis of the evidence presented by the first announcement of the discoveries of the Australopithecine fossils, there seemed at first sight to be several possible interpretations. The Australopithecinae might be nothing more than extinct varieties of ape closely akin to the chimpanzee and gorilla, but with certain modifications which in some minor respects show a spurious resemblance to the Hominidae. Secondly, they might have no special relationship to the gorilla and chimpanzee, but, nevertheless, represent a collateral group of anthropoid apes showing certain human characters developed as the result of a parallel evolution but not necessarily indicative of any real affinity with the Hominidae. Lastly, the Australopithecinae could be regarded as extinct hominoids which, while still at (or, at least, close to) the simian level in their cerebral development, were early representatives of the human branch of evolution and thus quite distinct from the Pongidae. This last interpretation has for a number of years been reiterated by Dart and Broom. On the other hand, other anatomists and paleontologists (particularly those who have not had the advantage of examining the original fossil material) have, either by direct statement or by implication, favoured one of the first two interpretations. As the result of his personal studies, the present writer has come to the firm conclusions (1) that the Australopithecinae have no special relationship to the recent anthropoid apes except so far as they are large hominoids of comparable size, and (2) that the human resemblances in the skull, dentition and limb bones are so numerous, detailed and intimate as virtually to preclude the introduction of the idea of ‘parallel evolution’ in order to explain them. In other words, there must be a real zoological relationship between the Australopithecinae and the Hominidae.
And so, the establishment came to see what is in hindsight obvious: the australopithecines were not merely the close relatives of modern apes, nor even such relatives that happened to acquire human-like characteristics through (rampant) convergent evolution. Rather, they were what Dart and Broom had maintained all along: extinct forms that were our close relatives – more closely related to us than to any living ape.
With the weight and stature of Le Gros Clark behind them, the evidence that Dart and Broom had painstakingly accumulated over decades finally gained scientific acceptance. Indeed, the weight of the evidence from the australopithecines – and Dubois’ earlier correct interpretation of Pithecanthropus (Homo) erectus now was seen for what it was: that the form of human body had evolved first, followed by an expansion in brain size. With this picture now in sharp focus, Piltdown stuck out like a sore thumb – and its days as an icon of human evolution were numbered.
Piltdown downfall
The end for Piltdown would be swift in coming, and Le Gros Clark would be one of the key figures in unmasking Piltdown for what it was. Already in the late 1940s a new dating test using fluorine had shown Piltdown’s skull to be of modern, not ancient age. Further analysis would reveal the extent of the deception – the skull of a modern human, the jaw of an orangutan, the teeth of a chimpanzee filed down, all stained with chemicals to look ancient. In the early 1950s the unraveled hoax would hit the popular press – a mystery solved, except for the identity of the forger. Though many individuals have been discussed as suspects in the following decades, the weight of evidence favors the original “discoverer” of Piltdown Man – Charles Dawson – who was the sole “discoverer” of the second Piltdown find, and, as it has been shown, forged many other lesser-known “discoveries” in hopes of fame and scientific standing. If he indeed was the forger, Piltdown was his crowning accomplishment, but he would take his secrets to an early grave in 1916 – without his desired knighthood or an appointment to the Royal Society. Regardless of who was responsible, Piltdown would cast a long shadow over the work of honest scientists such as Dubois and Dart – frustrating their attempts to advance what was a much more accurate view of human origins.
Lessons from Piltdown
Though Dubois and Dart faced an uphill battle, they did have a notable advantage: the nature of scientific advancement. As we noted right at the beginning of this series, evolution is a scientific theory – and as such, Dubois and Dart, with their genuine discoveries, needed only wait for additional work to move hominin paleontology onto a more solid scientific footing and provide a context for evaluating their efforts. The price they paid was not for being correct, but rather for being first, for which – even apart from fraud – there is often a price to pay in science. Piltdown would only prolong the problem, but in science, eventually the truth will out – for only the truth, or something close to it, is useful for making new, accurate predictions.
Next, we’ll summarize the current state of the evidence as it pertains to hominin evolution, and place the origins of our own species in that context.
In the early 1950s, with Piltdown unmasked, and the correct interpretation of Dart’s Australopithecus africanus and Dubois’ Pithecanthropus (Homo) erectus now accepted by the scientific community. Paleontologists now knew where to look (in Africa, as Darwin had suggested) and what to look for (species with human-like bodies and sub-human cranial capacities). In the following decades, many more finds would come to light –revealing the hominins to be a large group with numerous species. Rather than finding a “ladder” of species leading to humans, hominin paleontology would discover a branching tree of our close relatives.
As with all groups of fossil species, determining precise relationships is challenging. As we have pointed out previously in this series, the line between “species” is a fuzzy one, where separation is achieved slowly over time. Widely-separated species are easy to identify as distinct, but as one collects more and more data, the “gaps” start to be filled in, making distinctions more difficult. In a discipline like hominin paleontology this problem is somewhat heightened due to our intense interest in our own lineage – meaning that we have expended great effort to find as many hominin remains as possible. With an abundance of data, the picture becomes harder to resolve in fine detail even as the broad outlines become clearer. This leads to a phenomenon where some paleontologists favor delineating a large number of hominin fossil species, whereas others favor grouping hominin remains into fewer species. Informally, those who favor more species are known as “splitters,” and those who favor fewer species are known as “lumpers.” The fact that such a debate is even possible is testament to the robust, gradient-like data set we have available.
Hominin paleontology: a thumbnail sketch of the current evidence
In broad outline, the current state of hominin paleontology recognizes several large groupings, or grades, with, as we would expect, fuzzy boundaries. A consensus phylogeny representing current thinking is shown below, revealing the hominin family as the bushy tree that it is, and highlighting the abundance of evidence we have accumulated since the time of Dart and Broom:
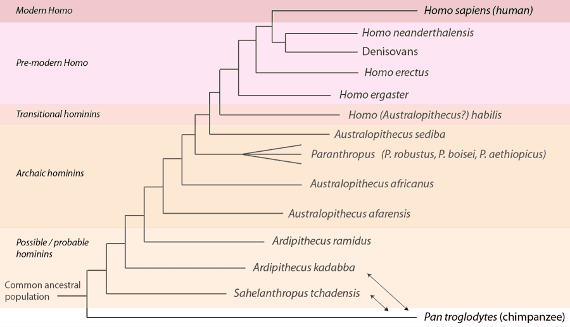
A hominin phylogeny based on current paleontological – and in some cases, genomic – evidence. Genome sequences are available for humans, Denisovans and Neanderthals. As we have previously discussed, Sahelanthropus tchadensis and Ardipithecus kadabbamay not be a hominins –they might be less closely related to humans than chimpanzees, in which case their positions would be switched with chimpanzees in the phylogeny (indicated with the double-headed arrows). Several hominin species are not shown. See text for details.
Working from the past towards the present, the first grouping we encounter includes species that we have discussed previously – ones that sit close to the human-chimpanzee last common ancestral population. This group, known as “possible and probable hominins” includes the “possible” hominin species Sahelanthropus tchadensis, Orrorin tugenensis, and Ardipithecus kadabba, but only one “probable” hominin, Ardipithecus ramidus. As we noted before, the evidence for bipedality is strongest for Ardipithecus ramidus. While “splitters” recognize these species as distinct, “lumpers” group them together as Ardipithecus ramidus sensu lato, where“sensu lato” indicates “in the broad sense.” In general this grouping exhibits a small braincase volume (under 350 cubic centimeters) and some hints of bipedality.
The second grouping, “archaic hominins” includes the Australopithecines such as Dart’s Australopithecus africanus and an earlier species, Australopithecus afarensis – the most famous example of which is nicknamed “Lucy.” Also included in this general group are the “robust” archaic hominins, the various species assigned to Paranthropus (such as Bloom’s Paranthropus robustus). Generally speaking, in this grouping we see growing evidence of occasional (i.e. facultative) bipedality and a further increase in braincase volume to a range encompassing 500cc. While splitters see numerous additional species in this grouping, lumpers recognize onlyA. afarensis, A. africanus, and one species encompassing all of Paranthropus.
The third grouping, “transitional hominins” contains the oldest-known members of Homo, such as Homo habilis. Underscoring their transitional nature, however, some scientists do not place this species within Homo, but rather describe it as an australopithecine, Australopithecus habilis. Braincase volume in habilis ranges between 500cc and about 700cc. A similar species, Homo rudolfensis, is recognized by splitters but grouped together as H. habilis sensu lato by lumpers. These species (or this species, as the case may be) shows increasing evidence of bipedalism, but not the full commitment to a bipedal lifestyle (i.e. obligate bipedalism) seen in later Homo. These groups also used stone tools of the Oldowan variety, a relatively simple stone tool technology named for the site of its first discovery.
The fourth grouping, “Pre-modern Homo” includes Homo ergaster and Dubois’ Homo erectus, the earliest Homo species that are fully bipedal. While splitters recognize ergaster and erectus as separate species, lumpers place them together as Homo erectus sensu lato. This grade also includes the more familiar Neanderthals as well as the Denisovans, species for which we have full genome sequences. Braincase volume in this grade starts at 700cc for early Homo ergaster/erectus, and extends to the volume seen in modern humans (about 1400cc) (and in fact exceeds average modern human volume in the Neanderthal lineage, where 1600cc skulls are found). Early in this grade, we see advancement to the more complex Acheulean stone tool technology develop.
The last grouping, “modern Homo”, contains only our own species, Homo sapiens, which enters the fossil record about 200,000 years ago. Splitters, however, have argued for an early subspecies, Homo sapiens idaltu, at about 160,000 years ago.
Ancestors, maybe: relatives, certainly
This diversity of remains both complicates the details of our evolution – in that we cannot be absolutely certain which (if any) of these groups are directly ancestral to our own species – and clarifies the overall picture, since these are our close relatives even if not our direct ancestors. Taken together, we can be confident that our lineage passed through these grades in turn – as an ardipithecine (or close relative), to an australopithecine (or close relative), and so on through the transitional hominins and on into pre-modern, and finally modern, Homo. Along the way, our lineage would expand in average braincase volume, acquire the use of ever more sophisticated tools, and, some 200,000 years ago, take our current form in Africa, prior to our expansion across the globe.
In the next post in this series, we’ll take up the story as our species expands out of Africa – and encounters the descendants of related lineages that left Africa before us.

About the author

Dennis Venema
Related resources
If you enjoyed this article, we recommend you check out the following resources:


Language of God
Sy Garte | Life’s Deeper Logic


Language of God

Language of God

Language of God