The Cambrian “Explosion”, Transitional Forms, and the Tree of Life
Many evolutionary critics have identified the Cambrian Explosion as a stumbling block to the theory of evolution. Keith Miller argues that the it is not as problematic as the opponents claim.
The most fundamental claim of biological evolution is that all living organisms represent the outer tips of a diversifying, upward-branching tree of life. The “tree of life” is an extremely powerful metaphor that captures the essence of evolution. Like the branches of a tree, as we trace individual lines of descent (lineages) back into the past (down the tree) they converge with other lineages toward their common ancestors. Similarly, these ancient lineages themselves converge with others back in time. Thus, all organisms, both living and extinct, are ultimately connected by an unbroken chain of descent with modification to a common ancestral trunk among single-celled organisms in the distant past.
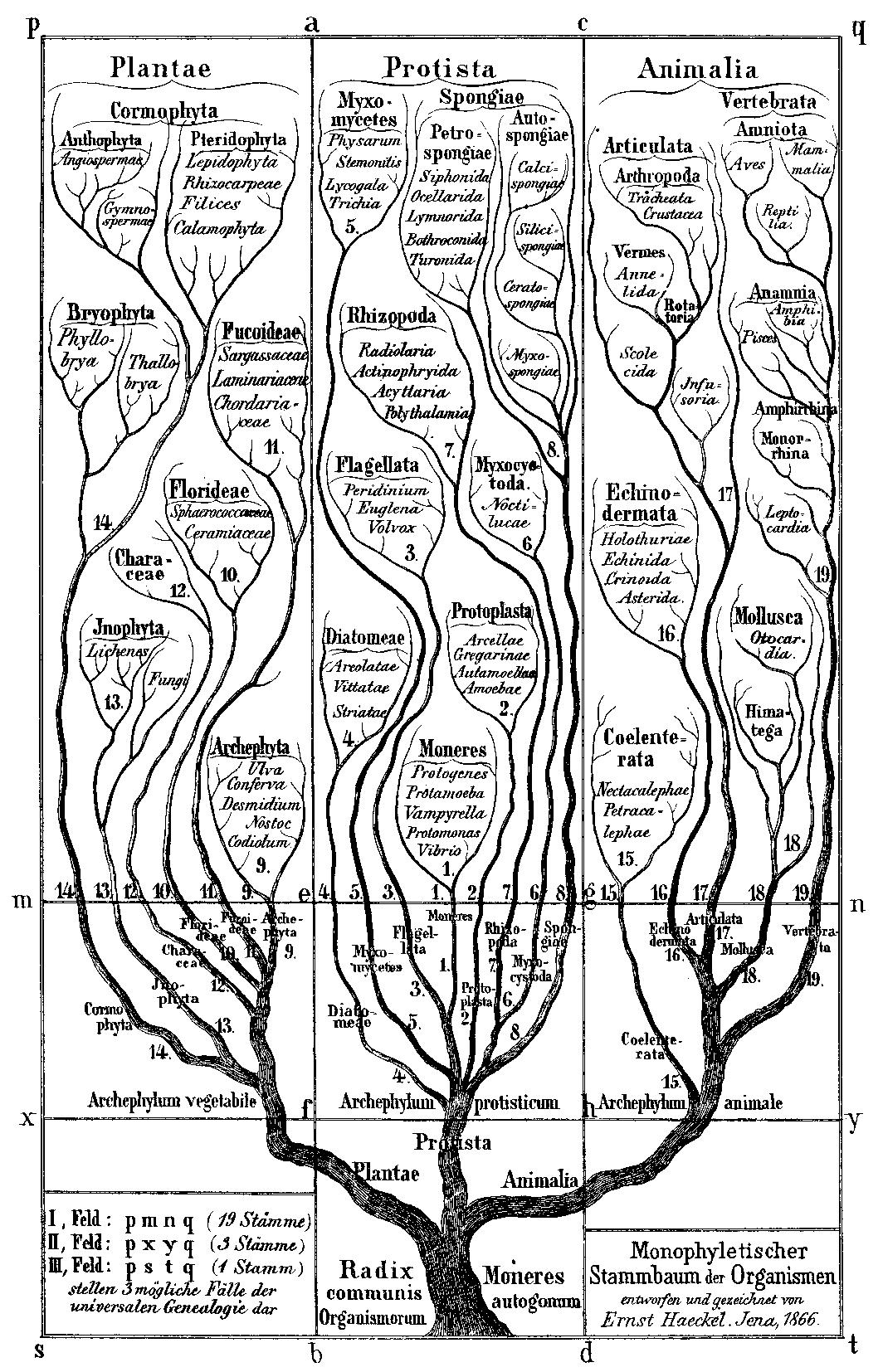
Click to enlarge
This tree metaphor applies as much to the emergence of the first representatives of the major groups of living invertebrates (such as annelids, snails, or arthropods) as it does to the first appearance and diversification of dinosaurs, birds, or mammals. This early diversification of invertebrates apparently occurred around the time of the Precambrian/Cambrian boundary over a time interval of a few tens of millions of years. This period of rapid evolutionary diversification has been called the “Cambrian Explosion.”
The Cambrian explosion has been the focus of extensive scientific study, discussion, and debate for decades, and is increasingly receiving attention in the popular media. It has also received considerable recent attention by evolution critics as posing challenges to evolution. These critics argue that the expected transitions between major invertebrate groups (phyla) are absent, and that the suddenness of their appearance in the fossil record demonstrates that evolutionary explanations are not viable.
What are some of the arguments of the evolution critics? John Morris of the ICR writes:
“If evolution is correct, the first life was quite simple, evolving more complexity over time. Yet the Cambrian Explosion of Life has revealed life’s complexity from the start, giving evolution a black eye. The vast array of complex life that appears in the lowest (or oldest) stratigraphic layer of rock, with no apparent ancestors, goes hard against evolutionary dogma. Evolution’s desperate attempt to fill this gap with more simple ancestral fossils has added more injury. …. Think of the magnitude of this problem from an evolutionary perspective. Many and varied forms of complex multi-celled life suddenly sprang into existence without any trace of less complex predecessors. There are numerous single-celled forms at lower stratigraphic levels, but these offer scant help in solving the mystery. Not one basic type or phyla of marine invertebrate is supported by an ancestral line between single-celled life and the participants in the Cambrian Explosion, nor are the basic phyla related to one another. How did evolution ever get started?”1
Intelligent design advocate Stephen Meyer and others have written:
“To say that the fauna of the Cambrian period appeared in a geologically sudden manner also implies the absence of clear transitional intermediates connecting the complex Cambrian animals with those simpler living forms found in lower strata. Indeed, in almost all cases, the body plans and structures present in Cambrian period animals have no clear morphological antecedents in earlier strata.2
And:
“A third feature of the Cambrian explosion (as well as the subsequent fossil record) bears mentioning. The major body plans that arise in the Cambrian period exhibit considerable morphological isolation from one another (or “disparity”) and then subsequent “stasis.” Though all Cambrian and subsequent animals fall clearly within one of a limited number of basic body plans, each of these body plans exhibits clear morphological differences (and thus disparity) from the others. The animal body plans (as represented in the fossil record) do not grade imperceptibly one into another, either at a specific time in geological history or over the course of geological history. Instead, the body plans of the animals characterizing the separate phyla maintain their distinctive morphological and organizational features and thus their isolation from one another, over time.”3
Are these critiques warranted? To what extent is the Cambrian explosion really problematic for the evolutionary picture of an unbroken tree of life extending back to the earliest life on Earth?
Geologic Time Scales: How big was the bang?
The relative rapidity of the diversification of invertebrates during the Cambrian “explosion” is set against the backdrop of the Earth’s geologic and biologic history. Geologic time is unfamiliar to most people, and its shear vastness is difficult to grasp.
Two lines of evidence impact our understanding of the duration of the animal diversification that led to the appearance of the major groups of living invertebrates. The first is the dating of critical strata within the geological timeline such as the Precambrian-Cambrian boundary and various important fossil-bearing horizons. The second is the time of appearance of the first widely recognized fossil representatives of the major living groups (phyla) of invertebrate animals. The latter is in considerable flux as new fossil discoveries are made.
Originally, the base of the Cambrian had been set at the earliest appearance of organisms with mineralized skeletons – particularly trilobites. However, a diverse collection of tiny mineralized plates, tubes and scales was discovered to lie below the earliest trilobites.4 This interval of “small shelly fossils” was designated the Tommotian. Because of the presence of even earlier tiny mineralized tubes and simple burrows, there was no internationally accepted definition for the boundary until 1994. At that time, the base of the Cambrian was placed at the first appearance of a particular collection of small fossil burrows characterized by Treptichnus pedum.
Until the early 1990’s the age of the Precambrian-Cambrian boundary was not tightly constrained, and was estimated to be about 575 million years ago. However, in 1993 new radiometric dates from close to the accepted Precambrian-Cambrian boundary revealed that it was significantly younger — about 544 million years.5 A more precise date of 542 ± 0.3 million years has recently been formally accepted by the International Commission on Stratigraphy. The basis for this date was the discovery that a sharp worldwide fall (or negative spike) in the abundance of the isotope carbon-13 was coincident with the Cambrian boundary as previously defined. In Oman, this isotopic marker also coincides with a volcanic ash layer that yielded the 542 million year date using uranium/lead radiometric methods.6 This horizon also marks the last occurrence of several fossils characteristic of the underling late Precambrian Ediacaran Period.7 Such extinction events are commonly used to subdivide the geologic time scale.
The earliest diverse fossil invertebrate communities of the Cambrian are represented by the Chengjiang, in China. These deposits are dated at 525-520 million years. The famous Burgess Shale is considerably younger, dating at about 505 million years, and the end of the Cambrian Period is set at 490 million years. The Cambrian Period thus lasted for 52 million years. To put this in perspective, the time elapsed since the extinction of the dinosaurs at the end of the Cretaceous has been 65 million years. The Cambrian was a very long period of time.
If the Cambrian explosion is understood to comprise the time from the base of the Cambrian to the Chengjiang fossil beds, then this period of diversification in animal body plans appears to have lasted about 20 million years. However, not all living animal phyla with a fossil record first appear within this time window. The colonial skeleton-bearing bryozoans, for example, are not known from the fossil record until the end of the Cambrian around 491 million years ago.8 More significantly, several living invertebrate phyla have a fossil record that extends into the late Neoproterozoic before the Cambrian. Sponges have been recognized as early as 580 million years, cnidarians are present among the Ediacaran animals at around 555 million years, and the stem groups for some other phyla were also likely part of the Ediacaran communities.
Defining the Cambrian “explosion” is not as straightforward as it might seem. Although there was clearly a major burst of evolutionary innovation and diversification in the first 20 million years or so of the Cambrian, this was preceded by an extended period of about 40 million years during which metazoans (multicellular animals) arose and attained critical levels of anatomical complexity. The Ediacaran saw the appearance of organisms with the fundamental features that would characterize the later Cambrian organisms (such as three tissue layers, and bilaterally symmetric bodies with a mouth and anus), as well as the first representatives of modern phyla. The base of the Cambrian is not marked by a sharp dramatic appearance of living phyla without Precambrian roots. It is a subjectively defined point in a continuum. The Cambrian “explosion” appears to have had a “long fuse.”
The procedure of classifying organisms is called taxonomy, and the general name for individual groups is “taxa.” Significantly, the first question that needs to be addressed is — What is a phylum? A phylum is often identified as a group of organisms sharing a basic “body plan,” a group united by a common organization of the body. However, phyla can be understood fundamentally, like all other taxonomic categories, as groupings of taxa that are more closely related to each other than to any other group.
The most widely accepted method for grouping organisms today is called cladistics. In cladistics all taxonomic groups are monophyletic, that is all of the members of the group are descended from a common ancestor that is the founding member of that taxon. A branch of the tree of life whose members all share the same ancestor is called a “clade” – thus the term cladistics. Closely-related taxa that do not share the same common ancestor are called “sister” taxa. These sister taxa commonly resemble each other more than the descendant relatives resemble the ancestors of their clade. As a result, placing these organisms into their correct monophyletic groups can be very difficult. Thus, organisms within a given phylum may bear close similarities to those from another closely-related sister phylum. In fact, the assignment of a given organism or fossil specimen to a phylum can be just as problematic as assignments to lower-ranked taxa such as classes, orders, families, etc.9
Further complicating the assignment of fossil organisms to phyla is that the anatomical characteristics that are used to define living phyla did not appear simultaneously, but were added over time. This has resulted in the distinction between “crown groups” and “stem groups” in the scientific literature (see figure above). A crown group is composed of all the living organisms assigned to that phylum, plus all the extinct organisms that were descended from the common ancestor of those living organisms. The stem group is composed of organisms more closely related to one living phylum than to another, but that do not possess all of the distinguishing characters of the crown group. It turns out that the organisms appearing in the early Cambrian are, with few exceptions, not crown groups but stem groups. That is, the complete suite of characters defining the living phyla had not yet appeared. Many crown groups actually do not appear in the fossil record until well after the Cambrian.10
The existence of stem groups provides a way to understand how the basic body plan of a living invertebrate could have been built up in steps. The major invertebrate groups are often portrayed by evolution critics as possessing anatomies that are both irreducible in organization and separated from other groups by unbridgeable gaps. No transitions could exist even in principle. This view is illustrated by the following comment by John Morris:
“Let’s suppose you want to find the forefathers of the clams, a prominent resident of the Cambrian Explosion, for instance. As you follow the fossil clues into ever “older” strata, what do you find? You find clams. The first or lowest occurrence of clams is abrupt or sudden. There are no ancestors that are not clams. An evolutionary lineage is impossible to discern, for clams have always been clams. Fossil clams are quite abundant, found all over the world in rocks of every age, and clams live today. Great variety among them abounds, but they are still clams. Variety does not speak to ancestry. The same is true of all animals found in the Cambrian Explosion. How can evolutionary scientists use the fossils as evidence of a common descent of all life?”11
The phylum Mollusca, to which clams belong, actually illustrates well how modern body plans could evolve from earlier stem groups. There is a well-documented series of transitional forms that extends from pre-mollusks (stem mollusks) through primitive early mollusks to the first unambiguous clams. The animals in this group gradually acquired the whole set of characteristics we now use to define “clam”. The earliest known mollusk-like organism is Kimberella (fig.1) from the late Neoproterozoic Ediacaran. It is a primitive organism that appears to lack several features characteristic of modern mollusks and is thus a considered a stem mollusk. The first likely “crown group” mollusks appear in the earliest Cambrian as part of the “small shelly fauna.” While recognizable as mollusks, many of these fossils belong either to sister groups or to stem groups of living classes. The earliest fossil bivalves (“clams”) are linked through a series of transitional forms to two of these extinct groups – the rostroconchs (fig. 2) and the cap-shaped helcionelloids (fig. 3). The hinged valves of clams appear to have evolved by the lateral compression of cap-shaped shells and then the thinning and loss of shell material along the hinge line.12 The characters that we use to identify “clams” did not appear as a complete package, but were acquired over time.

Some critics of evolution make much of the “top-down” versus the “bottom-up” pattern of appearance of higher taxa. That is, phylum-level diversity reaches its peak in the fossil record before class-level diversity, and the class-level diversity before that of orders, etc. These critics interpret this apparent “top-down” pattern as contrary to expectations from evolutionary theory. For example, Stephen Meyer and others have argued:
“Instead of showing a gradual bottom-up origin of the basic body plans, where smaller-scale diversification or speciation precedes the advent of large-scale morphological disparity, disparity precedes diversity. Indeed, the fossil record shows a “top-down” pattern in which morphological disparity between many separate body plans emerges suddenly and prior to the occurrence of species-level (or higher) diversification on those basic themes.”13
However, this pattern is an artifact, being generated by the way in which species are assigned to higher taxa. The classification system is hierarchical with species being grouped into ever larger and more inclusive categories. When this classification hierarchy is applied to a diversifying evolutionary tree, a “top-down” pattern will automatically result. Consider species belonging to a single evolving line of descent given genus-level status. This genus is then grouped with other closely related lines of descent into a family. The common ancestors of these genera are by definition included within that family. Those ancestors must logically be older than any of the other species within the family. Thus the family level taxon would appear in the fossil record before most of the genera included within it. Another way of looking at this is the fact that the first appearance of any higher taxon will be the same as the first appearance of the oldest lower taxon within the group. For example, a phylum must be as old as the oldest class it contains. Most phyla contain multiple classes, which in turn include multiple orders, and so forth. Thus, each higher taxon will appear as early as the first of the included lower taxa.
Additionally, higher taxonomic levels typically reflect more general aspects of the body plan. Thus, a poorly preserved specimen may be confidently assigned to a particular phylum, but not to any one class. Similarly, a primitive fossil might have the distinctive features of a particular phylum, but not be clearly assignable to any particular class because it is a transitional form — that is, a stem group or a sister group to a living class of organisms. Both of these factors would promote the earlier recognition of higher taxonomic categories than lower ones. The “top- down” pattern of taxa appearance is therefore entirely consistent with a branching tree of life.
There is one last bias in our reconstruction of the past that is generated by the process of assigning organisms to particular phyla. Because phyla are defined by particular anatomical character traits, they cannot be recognized in the fossil record until after those specific characters evolve. However, the splitting of the branch of the tree of life to which a phylum belongs may have occurred many millions of years previous to the evolution of those characters. The actual first appearance of a phylum thus occurs after significant anatomical evolution has occurred along that particular branch of the tree. Branching points in the tree of life will always be older than the named taxa.14
There are two opposite errors which need to be countered about the fossil record: 1) that it is so incomplete as to be of no value in interpreting patterns and trends in the history of life, and 2) that it is so good that we should expect a relatively complete record of the details of evolutionary transitions within all or most lineages.
What then is the quality of the fossil record? It can be confidently stated that only a very small fraction of the species that once lived on Earth have been preserved in the rock record and subsequently discovered and described by science.
There is an entire field of scientific research referred to as “taphonomy” — literally, “the study of death.” Taphonomic research includes investigating those processes active from the time of death of an organism until its final burial by sediment. These processes include decomposition, scavenging, mechanical destruction, transportation, and chemical dissolution and alteration. The ways in which the remains of organisms are subsequently mechanically and chemically altered after burial are also examined — including the various processes of fossilization. Burial and “fossilization” of an organism’s remains in no way guarantees its ultimate preservation as a fossil. Processes such as dissolution and recrystallization can remove all record of fossils from the rock. What we collect as fossils are thus the “lucky” organisms that have avoided the wide spectrum of destructive pre- and post-depositional processes arrayed against them.
Soft-bodied organisms, and organisms with non-mineralized skeletons have very little chance of preservation under most environmental conditions. Until the Cambrian nearly all organisms were soft-bodied, and even today the majority of species in marine communities are soft-bodied. The discovery of new soft-bodied fossil localities is always met with great enthusiasm. These localities typically turn up new species with unusual morphologies, and new higher taxa can be erected on the basis of a few specimens! Such localities are also erratically and widely spaced geographically and in geologic time.
Even those organisms with preservable hard parts are unlikely to be preserved under “normal” conditions. Studies of the fate of clam shells in shallow coastal waters reveal that shells are rapidly destroyed by scavenging, boring, chemical dissolution and breakage. Occasional burial during major storm events is one process that favors the incorporation of shells into the sedimentary record, and their ultimate preservation as fossils. Getting terrestrial vertebrate material into the fossil record is even more difficult. The terrestrial environment is a very destructive one: with decomposition and scavenging together with physical and chemical destruction by weathering.
The potential for fossil preservation varies dramatically from environment to environment. Preservation is enhanced under conditions that limit destructive physical and biological processes. Thus marine and fresh water environments with low oxygen levels, high salinities, or relatively high rates of sediment deposition favor preservation. Similarly, in some environments biochemical conditions can favor the early mineralization of skeletons and even soft tissues by a variety of compounds (eg. carbonate, silica, pyrite, and phosphate). The likelihood of preservation is thus highly variable. As a result, the fossil record is biased toward sampling the biota of certain types of environments, and against sampling the biota of others.
In addition to these preservational biases, the erosion, deformation and metamorphism of originally fossiliferous sedimentary rock have eliminated significant portions of the fossil record over geologic time. Furthermore, much of the fossil-bearing sedimentary record is hidden in the subsurface, or located in poorly accessible or little studied geographic areas. For these reasons, of those once-living species actually preserved in the fossil record, only a small portion have been discovered and described by science. However, there is also the promise of continued new and important discovery.
The forces arrayed against fossil preservation also guarantee that the earliest fossils known for a given animal group will always date to some time after that group first evolved. The fossil record always provides only minimum ages for the first appearance of organisms.
Because of the biases of the fossil record, the most abundant and geographically widespread species of hardpart-bearing organisms would tend to be best represented. Also, short-lived species that belonged to rapidly evolving lines of descent are less likely to be preserved than long-lived stable species. Because evolutionary change is probably most rapid within small isolated populations, a detailed species-by-species record of such evolutionary transitions is unlikely to be preserved. Furthermore, capturing such evolutionary events in the fossil record requires the fortuitous sampling of the particular geographic locality where the changes occurred.
Using the model of a branching tree of life, the expectation is for the preservation of isolated branches on an originally very bushy evolutionary tree. A few of these branches (lines of descent) would be fairly complete, while most are reconstructed with only very fragmentary evidence. As a result, the large-scale patterns of evolutionary history can generally be better discerned than the population-by-population or species-by-species transitions. Evolutionary trends over longer periods of time and across greater anatomical transitions can be followed by reconstructing the sequences in which anatomical features were acquired within an evolving branch of the tree of life.
A very important question is what organisms existed before the Cambrian “explosion.” Were there Precambrian precursors, or did the Cambrian explosion really happen in a biological vacuum? Many critics of evolution claim that the Precambrian is devoid of fossils that could represent body plans ancestral to those of the Cambrian invertebrates.
The words of Darwin are often cited as evidence of the seriousness of the problem for evolution.
“There is another and allied difficulty, which is much more serious, I allude to the manner in which species belonging to several of the main divisions of the animal kingdom suddenly appear in the lowest known fossiliferous rocks. Most of the arguments which have convinced me that all the existing species of the same group are descended from a single progenitor, apply with equal force to the earliest known species.”1
When Darwin published his model of descent with modification by means of natural selection, knowledge of the fossil record was in its infancy. In particular, the Precambrian and Early Cambrian fossil record was virtually unknown. Even the fossils of the now famous Burgess Shale and similar units were as yet undiscovered. After more than a century of paleontological work, the situation has changed dramatically. In keeping with evolutionary expectations, fossils are now known from the late Precambrian and early Cambrian that record several dramatic transitions in the history of life.
The presence of Late Precambrian animals was recognized in the 1950s and became widely publicized by the early 1970s. These are the famous Ediacaran fossils named for fossil-rich beds in the Ediacaran Hills of South Australia and now recognized at sites throughout the world. These organisms are typically preserved as impressions in sandstones and siltstones. Associated with these fossils are trails (Figure 1) and simple burrows of organisms that show a limited increase in complexity and diversity toward the Cambrian.
The record of life actually extends far beyond the Ediacaran fossils (~565-545 My) into the deep geologic past. Fossils of algae, protists, and bacteria (Figure 2) are present throughout much of the precambrian. The earliest convincing fossils of bacteria are recognized in rocks 3.5 billion years old, and chemical signatures point to the presence of life even earlier. Finely layered mounds (called stromatolites) produced by the activity of mat-building bacteria and algae appear at about this time and become relatively abundant by around 2.7 billion years ago. Evidence of eukaryotic algae, possessing membrane-bounded nuclei and internal organelles, dates to about 1500 million years ago, or earlier if chemical evidence is accepted. Multicellularity had appeared by 1000 million years ago in the form of diverse and relatively advanced seaweeds. The earliest fossils of metazoans (multi-celled animals) may be represented by simple disk-shaped fossils found in rocks 610-600 million years old.2
The earliest unambiguous indication of the rise of metazoan life is preserved in the spectacular phosphorite deposits of the Doushantuo Formation of China dating to at least 580 million years ago. Phosphate can preserve organisms and tissues in such great detail that individual cells can often be recognized. Where environmental conditions are ideal for this type of preservation, extraordinary fossil deposits may result. In the case of the Doushantuo, phosphatization has preserved not only a variety of algal remains, but also the cellular tissues of sponges and millimeter-sized tubes that might represent stem cnidarians.17 However, even more spectacular is the preservation of metazoan eggs and early embryos. These embryos (see Figure 3) are of uncertain affinities but may represent cnidarians or even bilaterians (animals with bilateral symmetry).18
The Ediacaran fossils provide the next window into the rise of metazoans. These fossil-bearing units span from about 575 million years to the base of the Cambrian, and are found in south Australia, Namibia, the White Sea coast of Russia, and Newfoundland. The enigmatic soft-bodied organisms were preserved as impressions, or molds, on the surfaces of sandstone and siltstone layers. These sediment layers accumulated in shallow-marine environments where the seafloor was covered by firm microbial algal mats. The microbial mats covering the seafloor appear to have been important in determining the lifestyles of the Ediacaran organisms, as well as their unique mode of preservation.5
Most soft-bodied impressions of the Ediacaran (or Vendian) can roughly be placed into three general groups — disks, fronds, and flat-bodied, bilaterally-symmetric forms. The biological affinity of these fossils is very difficult to determine and highly debated.6 Disks are the earliest appearing, and most common, Ediacaran fossils. They have often been identified as medusoids (“jellyfish”) but many appear to have been attached to the bottom, and none bear clear structures that would place them in a living group. Some do clearly possess tentacles around their margins suggesting a stem or sister group relationship to the cnidarians. Some sack-shaped fossils might even be stem anthozoans (the cnidarian group that includes anemones and corals).7
A few disk-shaped fossils may be related to other living phyla. One such form appears to be a sponge that might be assignable to the modern class of hexactinellids.8 Another is a small disk that has a raised center with five radial grooves that has been interpreted as a stem echinoderm (the phylum that includes modern starfish and sea urchins) that lacked the characteristic porous calcareous plates and other diagnostic features of true echinoderms.9
The frond-shaped forms include organisms that were attached to the bottom by a stalk, and others that appear to have been free lying. These fossils have also been assigned by some workers to a group of modern cnidarians (the “sea pens”) or to ctenophores. However, like the disks, the fronds are fairly diverse and some may be unrelated to living phyla.10 Others, although likely not able to be placed into a living cnidarian group, may be stem cnidarians, or even stem anthozoans. The discovery of better preserved fronds in the Cambrian that closely resemble some of the Ediacaran fossils would seem to support this interpretation.11
The bilaterally-symmetric forms of the Ediacaran are the most diverse and most enigmatic fossils of the late Precambrian. Some of these fossils may represent early experiments on the pathway to the living phyla.12 For example, Dickinsonia (Figure 4) and the similar Yorgia are fairly large flat highly-segmented forms that some workers have interpreted as annelids or stem annelids, while others have seen resemblances to other worm phyla or even chordates. These organisms do appear to have been able to move about the bottom as seen by associated crawling and resting traces. Even if not members of a living phylum, these organisms appear to at least be mobile bilateral metazoans (or bilaterians). Another bilateral form that has been the subject of much recent attention is Kimberella (Figure 5). This 555 million year old fossil has been interpreted as a stem mollusk.13 Scratch marks found associated with Kimberella indicate that it had some form of feeding structure (though probably not a true mollusk radula) that enabled it to graze the abundant algal mats. Other bilateral fossils have been interpreted to bear similarities to arthropods, although these interpretations are disputed.
An important, but less attention-getting, component of the Ediacaran fossil record is the presence of trace fossils such as trails, burrows and feeding traces. Except in the few cases mentioned above, there are no body fossils preserved of the organisms that made these traces. These traces tend to be small unbranched sediment-filled burrows that run horizontally along the sediment surface or under the microbial algal mats. Somewhat more complex burrows appear toward the base of the Cambrian including irregularly branching burrows and shallow vertical burrows.14 These traces are important because they point to the existence of small worm-like organisms that were probably feeding on and in the algal mats that covered extensive areas of the seafloor. The biological identity of these organisms is unknown, although they were clearly bilateria.
There is one more set of fossils that are known from the late Ediacaran (550-543 million years) that reveal yet another aspect of the metazoan diversity before the Cambrian. These fossils include tiny calcified or phosphatized tubes, cones and goblet-shaped structures that record the presence of animals capable of producing mineralized skeletons. They are commonly embedded within algal buildups that formed reef-like structures, and may be quite abundant.15 These algal-metazoan reefs foreshadow the later algal reefs of the Cambrian. The very peculiar cm-sized goblet-shaped Namacalathus (found as calcified fossils) lived attached to the algal mounds by stalks (Figure 6). Although the preserved shape of these fossils is consistent with that of cnidarians, their biology is uncertain.
The cone-in-cone structures of Cloudina (Figure 7) , and the more tubular Sinotubulites could have been produced by various types of worms such as serpulids. However, as with the trace fossils, the identity of the actual tube formers remains unknown. A significant observation of the Cloudina fossils is that many of them are perforated by borings. These borings provide the first clear evidence of predation before the Cambrian.
It is clear from the above discussion of the latest Precambrian, that the Cambrian explosion did not occur in a biological vacuum. Although many of the fossil specimens are enigmatic and difficult to classify, they nonetheless show significant biological diversity. Furthermore, at least a few living phyla had already appeared by the beginning of the Cambrian, and other forms likely represented stem groups related to later-evolving phyla.
One of the most important features of the Cambrian “explosion” was the very rapid diversification of organisms with shells, plates, and various other types of hard parts. A wide variety of soft-bodied organisms are also known from the Cambrian. Although some fossils can be assigned to living phyla, there are also specimens that appear to represent stem groups or intermediates between modern phyla, as well as specimens of unknown relationship. Representatives of several living classes and other lower taxonomic categories also appear in the Cambrian. A few deposits with exceptionally good preservation of fossils, such as the Burgess Shale in Canada, contribute to the wide range of taxa known from the Cambrian. Such deposits with exceptional preservation are known as Konservat-Lagerstätten (from the German “conservation deposits”). Similar deposits have since been found around the world in the Early to Middle Cambrian, notably the Early Cambrian Chengjiang fauna of China. Additionally, trace fossils become much more varied, complex, and abundant in the Cambrian, suggesting a newly widened range of animal activity.
Some of the very first fossils to appear near the base of the Cambrian are tiny skeletal plates, spines, tubes, and cap-shaped shells that have been called the “small shelly fossils.”1 Among these are the spicules of different groups of sponges, and the shells of the earliest known “crown group” mollusks and brachiopods. However, the biological identities of many of these tiny skeletal elements were completely unknown until fairly recently. Well-preserved complete fossils in the Chengjiang, and other fossil lagerstätten around the world, have revealed that some of these small shelly fossils were actually the spines and “armoring” of larger metazoans. More detailed analysis of other fossils has revealed that they may represent the stem groups of living phyla, rather than evolutionary dead ends.
The discovery of complete specimens from later in the early Cambrian has revealed that a variety of scales, plates and spines found among the “small shelly fossils” actually fit together and overlapped to cover the bodies of slug-like organisms.2 These organisms are the halkieriids and wiwaxiids. The halkieriids bore conical mollusk-like shells as well as calcareous structures similar to the chitinous bristles typical of polychaete annelid worms. The slightly younger Wiwaxia was covered in scale-like and spine-like structures even closer to those of the polychaetes, and also possessed a radula diagnostic of mollusks. These various unusual organisms bear resemblances to both mollusks and polychaete annelid worms, which are closely related phyla. Thus these organisms would appear to be positioned somewhere on the evolutionary tree near the branching point of the mollusks with the annelids.
Other cap-shaped fossils from the earliest Cambrian are the helcionelloids. These are interpreted as monoplacophoran-like crown group mollusks. As discussed earlier in the section on “Classifying Animals”, there is good fossil evidence of the transition from these primitive cap-shaped helcionelloids to the first bivalves. There are also likely fossil transitions from helcionelloids to the first gastropods.
Another important group of organisms represented by small plates in the early Cambrian are the lobopods. Lobopodians, until very recently an enigmatic group of strange fossils, were “caterpillar-like” organisms with fleshy lobed limbs and mineralized plates or spines running along their backs. They are similar to the living Onychophora, or velvet worms, but are considered a distinct group.3 The oldest known lobopodian bears certain similarities to a distinctive group of worms called the palaeoscolecid priapulids that also bore small plates or tubercles along their bodies.4 Lobopods may have been derived from these worms that also have an early Cambrian fossil record. Furthermore, the lobopods have become recognized as the critical link in reconstructing the assembly of the arthropod body plan. They have anatomical features in common with the arthropods, particularly with peculiar Cambrian stem arthropods such as Opabinia and Anomalocaris that are preserved in the younger Chengjiang and Burgess fossil beds. These later organisms possessed lobopod limbs but also had gill flaps along their bodies and jointed feeding appendages. Intermediates between lobopodians and the early stem group arthropods have also been discovered that possessed gills.5 Of even greater interest is the evidence available from the extraordinary preservation of muscle tissue (this paper is available online here) in a few of these transitional organisms. These specimens suggest a progression of steps in the transformation of internal anatomy from lobopodians to true arthropods.6
The tommotiids, a group of roughly conical-shaped shells composed of calcium phosphate, have until recently been one of the most enigmatic of the small shelly fossils. However, new discoveries of articulated specimens have shown that pairs of symmetrical skeletal elements fit together to form an open cone that was attached to the seafloor at the base. An opening at the base indicates the presence of a muscular attachment structure likely similar to the pedicle of brachiopods. The paired shells also have features similar to the tiny paterinids, crown group brachipods with calcium phosphate shells that also appear in the early Cambrian.7 These fossils therefore appear to represent stem brachiopods that were themselves derived from armored tubular filter feeders attached to the seafloor.
Following the appearance of the small shelly fossils, the diverse metazoan fossil communities of the Chengjiang in China are dated at around 525-520 million years, 20 million years after the beginning of the Cambrian. The exceptional preservation in these fossil beds is similar to that of the Burgess Shale deposits that are dated around 515-505 million years. These extraordinary fossil sites give us our best views into the composition of marine biological communities from this time, preserving both soft-bodied organisms and those with mineralized skeletons.8These beds contain abundant and diverse sponges and cnidarians, as well as priapulid worms, annelid worms, lobopods, stem mollusks such as Wiwaxia, and brachiopods. However, probably the most dramatic characteristic of the Chengjiang and Burgess type deposits is the abundance and diversity of arthropods.
Arthropods comprise 50% or more of all of the fossil specimens collected from these beds. These fossils include stem arthropods such as the anomalocarids, trilobites, which came to dominate the Paleozoic, and some species that appear to be crustaceans and chelicerates. However, most of the fossils belong to primitive stem groups that likely represent evolutionary experimentations after the appearance of true arthropods but before the rise of most living arthropod groups. In the Burgess Shale one such primitive species alone (Marrella) comprises a third of all fossil specimens. These fossils show unusual arrangements, and types, of appendages.
The chordates (that include vertebrates), hemichordates (that include the living “acorn worms“), and echinoderms (that include the living starfish and echinoids) are all deuterostomes and have the same pattern of early embryo development. Although the modern representatives of these phyla appear extremely different, they are actually closely-related branches on the tree of life, and are understood to have evolved from a common ancestor. Some rare, but very significant, specimens in the Chengjiang seem to be stem chordates and stem echinoderms, as well as specimens that have been interpreted as organisms close to the common ancestors of chordates and echinoderms. These rather simple Cambrian organisms possess the anatomical characteristics that would be expected in organisms that had acquired some but not all of the distinctive features of chordates or echinoderms.
A very primitive stem group of deuterostomes, called ventulicolians has recently been described that might represent the anatomy of organisms near the base of the deuterostome evolutionary branch that were ancestral to both the chordates and echinoderms.9 These soft-bodied organisms possessed segmentation and oval structures interpreted as gill slits, and a terminal mouth. Significantly, another group of primitive deuterostomes, called vetulocystids, bears similarities to the ventulicolians as well as to some of the bizarre early echinoderms. These organisms were likely anchored to the sediment and possessed an echinoderm-like mouth and respiratory openings.10 They may in fact represent organisms ancestral to the first echinoderms that were characterized by peculiar globular and asymmetrical shapes.
The most primitive group of chordates are the urochordates, or tunicates that have a sack-like adult body that filters seawater through pharyngeal slits. In their tadpole-like larval form, they possess stiff notochords (a structure diagnostic of chordates) that is lost in the adult form. A likely tunicate has been described from the Chengjiang.11 Another group of primitive chordates are the cephalochordates (represented today by the lancelets that possess a notochord as adults, pharyngeal slits, and muscles arranged in parallel bundles. Some fossils have been interpreted as stem cephalochordates.12 Lastly, and of particular interest, is a fossil that may be a stem vertebrate.13 Haikouichthys, in addition to a notochord, gill pouches and muscle bundles, also appears to have had some structures characteristic of vertebrates. These vertebrate features include a cavity surrounding the heart, a dorsal fin, and cartilage around the head and as a series of elements along the notochord. The Chengjiang thus includes fossil specimens that occupy several significant transitional positions from primitive deuterostomes, to stem echinoderms and stem chordates.
The fossils of the Cambrian “explosion” were indeed diverse and included organisms that can be assigned to a number of living phyla. As we have seen, these fossil organisms were also largely representative of stem groups that possessed some, but not all, of the diagnostic features that define the major groups of living organisms. The body plans of phyla were assembled piecemeal. Furthermore, important transitional steps between living phyla and their common ancestors are also preserved. These include: the rise of mollusks from their common ancestor with the annelids, the evolution of arthropods from lobopods, the likely evolution of brachiopods from tommotids, and the rise of chordates and echinoderms from early deuterostomes. While the picture is far from complete, the spectacular fossil discoveries from the early and middle Cambrian strongly support the conclusion that the major branches of the animal tree of life are joined to a common metazoan (multi-cellular animal) trunk.
Numerous hypotheses exist for the geologically rapid diversification of invertebrates in the Cambrian, proposing various key evolutionary innovations or environmental triggers. Critical levels of ecological or behavioral complexity may also have stimulated diversification. At the molecular level, organisms may have reached a key threshold of genetic organization or evolved a key gene.
A number of important environmental changes occurred in the late Precambrian and in the early Cambrian that likely had important consequences for the early evolution of metazoans. Near the end of the Precambrian there were several episodes of nearly global glaciation in which sea ice and continental glaciation extended to the equatorial regions. The last of these “snowball Earth” episodes was about 635 million years ago.1 This time just precedes the earliest fossil evidence of metazoans. The major changes in ocean temperature and chemistry associated with the transition from a snowball Earth to a greenhouse world would likely have had profound effects on life. In particular, isotopic data indicates that the oceans became increasingly oxygenated after the end of the last of the global glaciations.2 Higher oxygen levels would have been critical for aerobic respiration and the evolution of larger body sizes.
The advent of mineralized hard parts was an important part of the Cambrian “explosion.” The ability of organisms to secrete hard parts had important consequences for both metazoan evolution, and for the preservability of these organisms in the fossil record. Much of the rapid increase in fossil diversity during the early Cambrian is among organisms with resistant hard parts. Changes in seawater chemistry may have played an important role in permitting or stimulating mineral precipitation by marine organisms. With the right concentrations of certain ions, normal physiological processes, such as respiration or photosynthesis, may cause precipitation. Such biomineralization could then be modified through natural selection. In addition, hard parts represent a handy way to store useful ions, or remove toxic ones. Carbonate and phosphate ions, present in most skeletons, are also good buffers against pH changes. Recent work on seawater chemistry during the latest Proterozoic and early Cambrian has indicated a major change in calcium ion concentrations between 544 and 515 million years.3 This time interval coincides with the onset of widespread biomineralization in the fossil record.
The rise of hard parts would likely have had important behavioral consequences. Hard skeletons provide firm attachments for muscles, enabling various activities and motions not otherwise possible, and skeletons would have helped to support larger and more complex bodies. Hard parts also would provide a protective armor against predators, and evidence for predation is found almost as early as the first skeletal elements appear in the fossil record. Predator-prey interactions seem particularly effective at producing an evolutionary escalation, with the prey evolving defenses and the predator evolving ways to overcome them. Animals with mineralized armor would promote selection for harder jaws and claws in the predators. The more effective predators would in turn increase selective pressure for more resistant skeletons in the prey.
Changes in animal behavior can also change the physical environment. A major environmental change in the early Cambrian came as a result of increased complexity and intensity of bioturbation (burrowing, digging, or other moving and mixing of the sediment by organisms). Burrowing can be a response to escaping predation or seeking out food resources. These evolving behaviors also disrupted the existing seafloor habitat. For much of the Precambrian and into the early Cambrian, microbial and algal mats largely covered the seafloor. These mats provided a stable base for sessile animals and kept mud out of the water, making it easy for filter feeders to obtain relatively high amounts of food and low amounts of sediment. The advent of algal grazers, extensive burrowing and other bioturbation disrupted these mats. This created problems for animals adapted to the old seafloor pattern, but provided a new habitat of muddy seafloors.4 Additionally, the constant burrowing unearthed buried nutrients, making them accessible to animals at the surface of the sediment.
Available food resources and ecological roles were also altered with the appearance of planktonic or swimming metazoans in the early Cambrian. Prior to the Early Cambrian, there is no evidence for macroscopic zooplankton or swimming animals. However, in the Cambrian several actively swimming, plankton-feeding animals appeared. At the same time, many kinds of planktonic algae became extinct and the surviving forms were much smaller. Evolution of swimming and plankton-feeding ability leads to the diversification of plankton feeders, but it also affects the bottom-dwelling organisms.5 Both the fecal material and the carcasses of these animals would have fallen to the bottom, moving large quantities of nutrients from the water column, where they were previously inaccessible to animals, to the sea floor. Even today, most of the nutrients in the deep sea come from these sources.
This brief survey of possible factors in the Cambrian explosion illustrates how ocean chemistry, environment, ecology and animal behavior are complexly intertwined. Complex positive and negative feedbacks make it very difficult to tease out which factor was most critical to the rapid diversification of metazoan life at the end of the Precambrian and early Cambrian. However, evidence from multiple sources strongly suggests that several significant changes in the world’s ocean environment conspired to light the fuse of evolutionary innovation.
Conclusions
Given our current, and continually growing, knowledge of the deep past, it is increasingly clear that the rise of multicellular animals is not an impenetrable mystery. While there is much that is not known, and will never be known, there is also much that has been discovered, and much excitement for what will yet be learned. The animals of the Cambrian did not appear in all their modern complexity out of a void, but rather provide pointers to their common ancestry. Despite the claims of evolution skeptics, the fossil record provides multiple examples of organisms displaying transitional anatomies. The anatomical characters that define the body plans of the major living animal phyla, can be seen to have been acquired piecemeal during the early evolution of the metazoa. Just as with all other taxonomic groups (e.g. classes, orders, families, genera, species), the divisions between phyla break down as we move closer to their times of origin from common ancestors. The tree of life continues to stand tall.
About the author

Keith Miller
Related resources
If you enjoyed this article, we recommend you check out the following resources:

Language of God
Sy Garte | Life’s Deeper Logic


Language of God

Language of God

Language of God

Language of God